Zoology
The Speed of the Nerve Impulse
Hermann von HELMHOLTZ. From Messungen ueber den xeitlichen verlauf der zuckung animalischer muskeln und die fortp flanzungsgeschwindigkeit der reizung in den nerven, in Archiv fuer anatomie, physiologie, und wissenschaftliche medicin herausgegeben von Dr. Johannes Mueller, p. 276, 1850; and from Messungen ueber fortp flanzungsgeschwindigkeit der reizen in den nerven, ibid., p. 199, 1852; both tr. by M. and V. Hamburger and T. S. Hall for this volume.
The duration of the twitch of an animal muscle is ordinarily only a small fraction of a second, except for a longer lasting, weak after-effect. Since our senses are not capable of immediate perception of single time elements of such short duration, we must use more artificial methods to observe and measure them. Two of these especially are to be considered here. In the first, the events whose time intervals one wishes to find out are recorded by a suitable mechanism on a surface which moves with even speed. The time intervals appear on it as proportional space differences and can be measured by the latter. Ludwig has already used this method for physiological purposes in order to show the fluctuations of blood pressure in the arteries and of atmospheric pressure in the pleural cavity. The other, essentially different, method of measuring time is the one proposed by Pouillet. The duration is here measured by the effect which a force of known intensity has produced during this interval. Pouillet has a galvanic current act on a resting magnet. The beginning and end of the current correspond exactly to the beginning and end of the interval to be measured; the magnitude of arc of the excursions which the magnet performs is, then, proportional to the duration to be measured. ...
The foundation of Pouillet’s method for measuring small time intervals is as follows: the time during which a galvanic current of known intensity from a coil has affected a magnet can be calculated exactly from its changed movement. Up to the present, one cannot anticipate a lower limit of time divisions measurable in this way, since one can increase at will the intensity of the acting current and the magnitude of its effect on the magnet by increasing the electromotor ceils and the windings on the coil. But a limitation is imposed in the application of this procedure; namely, one must know how to cause the beginning and end of the supposed current, which from now on we shall call the time-measuring one, to coincide exactly with the beginning and end of the mechanical process the duration of which is to be measured. In the experiments to be described here the time-measuring current started at the moment when an instantaneous electric shock passed through the muscle or its nerve, and stopped when the circuit within which it circulated was interrupted by the contraction of the muscle. At the same time one could determine exactly the tension which the muscle had to develop in order to be able to separate the conductive metals from each other. The duration of the time-measuring current to be calculated is therefore identical with the time which elapses between the stimulation of the muscle, or of its nerve, and the moment at which its tension reaches a certain magnitude. ...
[There follows a detailed description of an apparatus in which the nerve-muscle preparation is suspended and weighed down by a weight, with slight stretch being accomplished by overweight. Electrodes for stimulation by either galvanic or induction current could be placed at any point on the nerve. The time-measuring current is interrupted when the tension of the muscle, following stimulation, exceeds the sum of weight and overweight. This is measurable.—Ed.]
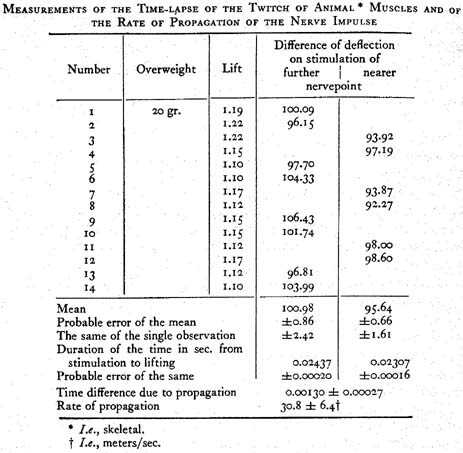
In making measurements of the time which elapses between the stimulation of the nerve and the lifting of the overweight by the muscle, one finds that the time depends upon the point on the nerve at which one applies the electrical shock; the time is the longer, the longer the portion of the nerve between the stimulated point and the muscle. The experiment ... can be repeated any number of times, by placing two of the four conducting wires, about two to three lines apart, on the nerve close to where it enters the muscle, and the two others, just as far apart, on the pelvic part of the nerve. I found it to be of advantage to move this second place not quite to the transected end of the sciatic plexus, but approximately to the place where the strands of this plexus combine to form the trunk of the sciatic nerve, because the extreme cut ends become inefficient relatively fast. Depending on whether one connects the first or the second pair of the leads with the induction coil, either the nerve point closer to the muscle, or the more distant one, will be affected by the current. Comparative measurements, which incidentally are carried out like those previously discussed, prove that the deflections of the magnet by the time-measuring current are on the average from 5–7 dial parts larger when the more distant point of the nerve is stimulated than the one closer to the muscle.
Apparently this difference cannot be caused by any of the formerly discussed sources of error, which are based on the mechanical and electrical occurrences in our measuring procedure, because all of these affect the experiments involving stimulation of the distant or near nerve point equally. Rather occurrences inside of the nerve itself must be the cause. ...
We must ... make sure that the intensity of stimulation is the same at both places. If this is so, then experiments show that whatever places on the nerve are stimulated, corresponding energy stages will follow each other at like time intervals, but the time between each of these energy stages (and the stimulation) is larger by a definite amount, as the stimulated spot is further away from the muscle. Therefore, if we express by curves the rise and fall of energy for two different nerve points, then the curve corresponding to the stimulation of the more distant point is congruent with the other, but between its starting point and the point corresponding to the moment of stimulation, there lies a larger part of the abscissa. From the nature of the time lapse which the muscle exhibits following stimulation we can draw conclusions concerning the course of the corresponding processes in the nerve which are mostly still unknown. ... Now, since duration and strength of the stimulating electric current are exactly the same in both stimulated places the retardation of the effect must be due to the fact that a certain time elapses until it has spread from the more distant spot to the muscle. These experiments, therefore, enable us to find out the rate of propagation of the impulse1 in the motor nerves of the frog, provided that we understand by impulse2 those processes in the nerve, which develop in it as a result of an external stimulus.2
As long as the physiologists thought that nerve action could be ascribed to the propagation of an imponderable or psychic principle, it would have appeared incredible that the speed of this current should be measurable within the short distances of the animal body. At present we know from the investigations on the electromotor properties of nerves by Du Bois-Reymond, that the activity by which the conduction of an impulse is mediated is at least closely associated with, perhaps even essentially caused by, a changed arrangement of their material molecules. Accordingly, the conduction in the nerve would belong to the group of propagated molecular effects of ponderable bodies, to which, e.g., belongs sound conduction in air and in elastic substances or the discharge of a tube filled with an explosive mixture. Under these circumstances, it is no longer surprising to see that the rate of conduction is not only measurable but as we shall see, even very moderate. Incidentally, the impossibility of observing time intervals of this kind in the daily perceptions of our own body, or in physiological experiments on muscle twitches must not surprise us, since the intervals which we may be sure that we observe between sensations involving the nerve fibers of our different sense organs are not much smaller than a second. One will recall that the most experienced astronomers differ by a full second in the comparative observation of visual and acoustic perceptions. ...
[There follows a description of the measures taken to eliminate differences in the degree of contraction when the near vs. the far end of the nerve is stimulated. It is reported that the far end of the nerve is the first to show a decrease of sensitivity following repeated stimulation.—Ed.]
From the greater number of my experimental series, all of which gave the same result, with more or less exactness, I shall present herewith those which seem to be the most reliable on account of their extent or the correspondence of their single observations. For stimulation, we have invariably used currents which brought about maximum excitation. This was controlled by simultaneously observed elevations expressed in millimeters.
The series are arranged according to different plans. In some of them, all observations are made with the same or only two different overweights, in order to get as extensive figures as possible for the calculation of the essential time interval. For these I have calculated the means of the time-lapse between stimulation and muscle reaction for both points on the nerve, the difference between these means which corresponds to the rate of conduction in the nerve, and finally, in order to evaluate their exactness, the probable errors of all these values according to the rules of probability.
In other experimental series, the overweights have been exchanged as often as possible in order to prove that the delay is the same, for different degrees of muscle energy, provided one stimulates from the more distant point on the nerve, but the form of the energy increase is not altered. Obviously, the few experiments made with each overweight cannot furnish such exact values for differences due to nerve conduction as would longer series; therefore the individual means for these differences often vary considerably. However, the larger and smaller values are distributed entirely irregularly, and those for different overweights do not differ more from each other than those for the same overweight in successive observations. It follows that the magnitude of the difference does not depend noticeably on the amount of overweight, as is so definitely the case when the deflections of the magnet increase by decrease of the stimulation.
Finally, the rate of propagation of the nerve impulse was calculated after each experimental series. To do this, one must know the length of the traversed nerve piece, that is, the distance between the terminals at the two stimulated nerve places closest to the muscle. Unfortunately this length is very uncertain on account of the great extensibility of the nerve. If the nerve is not stretched, its fibers are bent in an undulating fashion; in order to measure its length I have always stretched it to such an extent that the transverse satin-like striations of its surface disappeared, on the assumption that the fibers would then run approximately straight. But a few millimeters are then always left to one’s own discretion. Incidentally, it would not yet pay to devise an improved measuring technique since the inaccuracies of the time measurements are relatively much greater than those of the length measurements. Therefore it is not surprising that the established values of the rate of conduction still differ considerably from each other. ...
[The following is part of one of the twenty experimental series reported in the paper—Ed.]
SERIES X.
Done on December 29 with the muscles of a frog kept for four months. Through the more distant point on the nerve is sent a stronger current, generated with the coils touching each other, and through the nearer point a weaker current with a distance between the coils of 2½ cm. After each two observations the muscle is reset.
A. Right muscle—nerve length 40 mm., deflection before 116.09, after 112.45, mean 114.27.
The values found for the rate of propagation between 11 and 21ºC. are therefore:
a) from series IX, X, and XI.
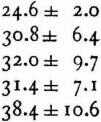
From these one finds by the method of least squares as most probable mean: 26.4 [meters/sec.].
b) from series XlI, XIII, and XIV.
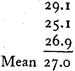
Finally, I summarize the results of the present investigations:
1) If animal (skeletal) muscle, or its nerve, is stimulated by a momentary electric shock, a short time passes during which its elastic tension does not change noticeably; then it gradually rises to a maximum, and just as gradually falls again. The contraction of animal muscle differs from that which occurs in organic (visceral), nonrythmically reacting muscle, after a relatively short stimulation, only in that its single phases pass much more rapidly.
If two different points of a motor nerve are stimulated by a momentary stimulus and if the magnitude of the stimulation is the same for both, then the time-lapse of the subsequent muscle twitch, is also the same; however, if the more distant point on the nerve has been stimulated, all of the muscle twitch stages occur later by an equal amount. From this, we conclude that the conduction of the nerve impulse to the muscle requires a measurable time. ... [In addition to Pouillet’s method, the graphic method was used. The results of these experiments are reported in the second paper, of which the essential portion follows.—Ed.]
In the first series of my investigations on the time relations of muscle and nerve activity I have proved by the electromagnetic method of measuring time, that the mechanical reactions of the muscle, following a nerve stimulation, set in later if the excitation has to pass a longer portion of the nerve before getting to the muscle. The method mentioned offers, in fact, the best guarantee where safe execution of exact measurements is desired, but it has the great disadvantage of yielding the said result only after extensive and tedious series of experiments, which on account of their long duration require an especially favorable condition of the frog preparation. The other graphic method of measuring time, the application Of which has been mentioned before, is essentially one in which the muscle during twitching records the magnitudes of its contractions on a moving surface; this promised a much simpler and easier demonstration of the rate of propagation in the nerves, and, since this seemed to me sufficiently important, I undertook to follow up the matter in this way, and I was perfectly successful.
The procedure of the experiments, I have already briefly indicated in the previous paper. A pen which is raised by the twitching muscle draws a curve on a surface moving with uniform speed, the vertical coordinates of the curve are proportionate to the contractions of the muscle, the horizontal ones proportionate to the time. As a starting point of this curve we shall fix that point which corresponds to the moment of stimulation of the muscle or its nerve. Now, if we arrange for two curves to be drawn in succession, and if we take care that at the moment of stimulation the pen occupies always exactly the same point on the surface, then both curves will have the same starting point, and from the congruence or noncongruence of their individual parts one can observe whether or not the different stages of the mechanical muscle response have occurred, in both instances, at the same or a later time after stimulation. ...
If the animal parts are rather vigorous and fresh, then the shapes of the double curves are all alike, at whatever nerve spot one may start the stimulation. Then, each drawing consists of two curves of congruent shape which are shifted in a horizontal direction with respect to each other by a certain amount as in Fig. II, such that the curve which has been drawn upon stimulation of the nearer nerve spot, is also nearer to the starting point of stimulation. The curve adefg corresponds to the stimulation of the nearer nerve point,
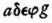
to the one of the more distant nerve point. ...
When we look at the double curve Fig. II it is evident that both of the muscle twitches recorded have been entirely identical as to strength, duration, and course of the different stages of contraction except that the one has
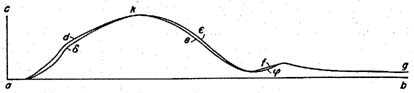 FIG. II.
FIG. II.
started later after stimulation than the other one. Now, since the arrangement of the apparatus and the mechanical forces of the muscle have been exactly the same, the delay of the reaction in one instance can only have been derived from the longer time of propagation in the nerve. ...
It is the great advantage of the described method that one can recognize immediately in each single drawing from the shape of the two curves whether the muscle has worked uniformly in both instances, whereas this fact could be deduced by the electromagnetic method of time measurement only from a long series of single experiments. As to the absolute value of the rate of propagation, the horizontal distances of the two curves cannot be measured with great accuracy; nevertheless, the values of that rate are about the same as in the former method. For instance, the horizontal distance in Fig. II is about I mm, the circumference of the cylinder corresponding to
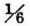
sec, is 85.7 mm, therefore the length of the abscissae is 514.2 mm per second. One mm corresponds therefore to 1/514.2 sec. The length of thè nerve involved in propagation was 53 mm, from which follows a rate of propagation of 27.25 m per sec. The most probable value from previous experiments was 26.4 m.
3
*Gesammelte Abhandlungen, 1877, Bd. ii. S. 319.
1 Helmholtz uses "Reizung" three times here, the distinction between stimulus and impulse being recognized in fact but not in words.
2 See footnote 1.
3 The modern figures for rate in frog gastrocnemius and sciatic nerve is 30–43 meters/sec. for coarse fibers, 16 meters/sec. for thinnest fibers.